O bomba de potássio de sódio É uma estrutura proteica incluída em um conjunto mais amplo de moléculas presentes em muitas membranas celulares, responsáveis pelo transporte ativo de íons ou outras pequenas moléculas contra seus gradientes de concentração. Eles usam a energia liberada pela hidrólise do ATP e por isso são chamados genericamente de ATPases..
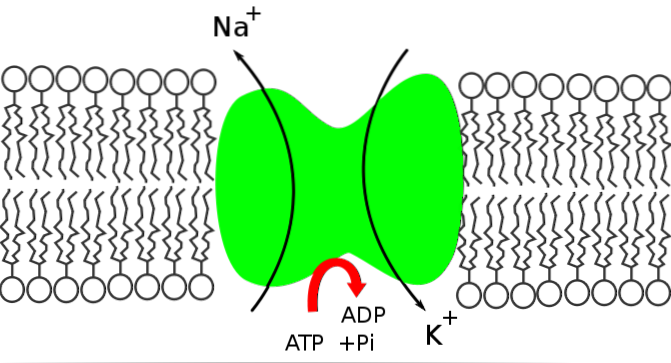
A bomba de sódio e potássio é uma Na + / K + ATPase porque libera a energia contida na molécula de ATP para mover o sódio de dentro para fora da célula, enquanto introduz potássio.
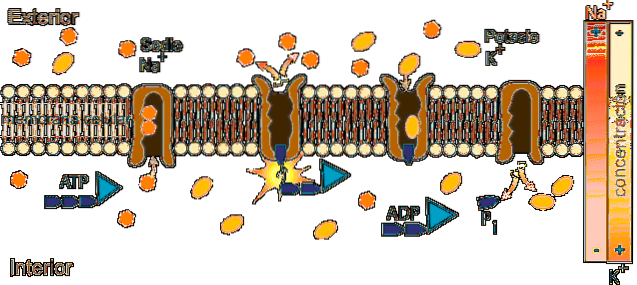
Dentro da célula, o sódio é menos concentrado (12 mEq / L) do que fora (142 mEq / L), enquanto o potássio está mais concentrado fora (4 mEq / L) do que dentro (140 mEq / L).
As bombas ATPase são classificadas em três grandes grupos:
A bomba de sódio e potássio (Na + / K + ATPase) pertence ao grupo das bombas iônicas da classe P e foi descoberta em 1957 por Jens Skou, um pesquisador dinamarquês, quando estudava o mecanismo de ação dos anestésicos nos nervos de um caranguejo. (Carcinus maenas); trabalho pelo qual recebeu o Prêmio Nobel de Química em 1997.
Índice do artigo
A bomba de sódio e potássio é uma enzima que do ponto de vista de sua estrutura quaternária é formada por 2 subunidades proteicas do tipo alfa (α) e duas do tipo beta (β)..
É, portanto, um tetrâmero do tipo α2β2, cujas subunidades são proteínas integrais de membrana, ou seja, atravessam a bicamada lipídica e possuem domínios intra e extracitosólicos..
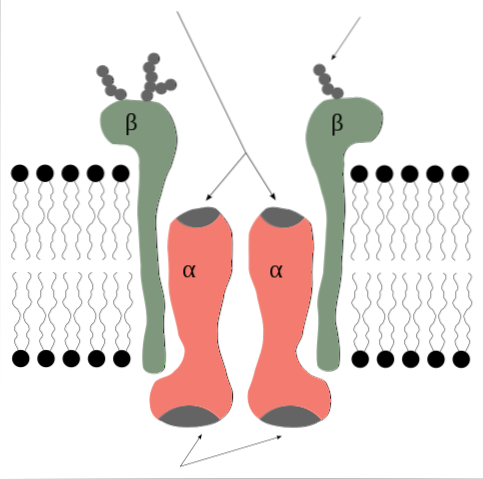
As subunidades α são aquelas que contêm os sítios de ligação do ATP e dos íons Na + e K + e representam o componente catalítico da enzima e aquele que exerce a função de bomba propriamente dita..
As subunidades α são grandes polipeptídeos, com peso molecular de 120 kDa, 10 segmentos transmembrana e com suas extremidades N- e C-terminais localizadas no lado citosólico..
No lado intracelular, apresentam sítios de ligação para ATP e Na +, além de um resíduo de aspartato na posição 376 que representa o sítio que sofre o processo de fosforilação durante a ativação da bomba..
O sítio de ligação para K + parece estar no lado extracelular.
As subunidades β não parecem ter uma participação direta na função de bombeamento, mas na sua ausência essa função não ocorre..
As subunidades β têm um peso molecular de cerca de 55 kDa cada e são glicoproteínas com um único domínio transmembranar cujos resíduos de carboidratos são inseridos na região extracelular..
Eles parecem ser necessários no retículo endoplasmático, onde contribuiriam para o dobramento adequado das subunidades α e, então, ao nível da membrana, para estabilizar o complexo..
Ambos os tipos de subunidades são heterogêneas e as isoformas α1, α2 e α3 foram descritas para uma, e β1, β2 e β3 para a outra. Α1 é encontrado nas membranas da maioria das células, enquanto α2 está presente no músculo, coração, tecido adiposo e cérebro e α3 no coração e cérebro.
A isoforma β1 é aquela com distribuição mais difusa, embora esteja ausente em alguns tecidos, como as células vestibulares do ouvido interno e as células musculares glicolíticas de resposta rápida. O último contém apenas β2.
As diferentes estruturas das subunidades que compõem a bomba Na + / K + nos diferentes tecidos podem ser devidas a especializações funcionais ainda não elucidadas..
Para qualquer momento considerado, a membrana plasmática constitui um limite de separação entre o compartimento correspondente ao interior de uma célula e aquele que representa o fluido extracelular no qual está imersa..
Ambos os compartimentos têm uma composição que pode ser qualitativamente diferente, uma vez que existem substâncias no interior das células que não se encontram fora delas e o líquido extracelular contém substâncias que não estão presentes intracelularmente..
As substâncias que estão presentes em ambos os compartimentos podem ser encontradas em diferentes concentrações e essas diferenças podem ter significado fisiológico. Esse é o caso de muitos íons.
A bomba Na + / K + desempenha papel fundamental na manutenção da homeostase intracelular por meio do controle das concentrações de íons sódio e potássio. Esta manutenção da homeostase é alcançada graças a:
Na + tem uma concentração extracelular de cerca de 142 mEq / L, enquanto sua concentração intracelular é de apenas 12 mEq / L; K +, por outro lado, está mais concentrado dentro da célula (140 mEq / L) do que fora dela (4 mEq / L).
Embora a carga elétrica desses íons não permita sua passagem através da membrana, existem canais iônicos que o permitem (seletivamente), que promovem o movimento se as forças que normalmente movem esses íons também estiverem presentes..
Agora, essas diferenças de concentração são de grande importância no preservação da homeostase do organismo e deve ser mantido em uma espécie de equilíbrio que, se perdido, produziria alterações orgânicas importantes.
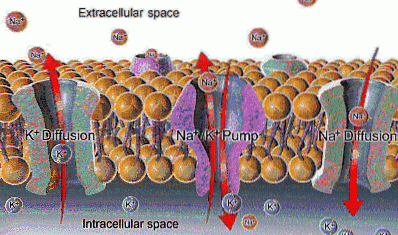
A função da bomba Na + / K + permite a extração do sódio que entrou por difusão através de canais ou outras vias de transporte e a reintrodução do potássio que se difundiu, permitindo a conservação das concentrações intra e extracelulares desses íons.
O mecanismo de ação da ATPase Na + / K + consiste em um ciclo catalítico que envolve reações de transferência de um grupo fosforila (Pi) e mudanças conformacionais da enzima que vai de um estado E1 para um estado E2 e vice-versa..
A operação requer a presença de ATP e Na + dentro da célula e K + no fluido extracelular.
O ciclo começa no estado de conformação E1 da enzima, no qual existem 3 sítios citosólicos de ligação ao Na + e alta afinidade (Km de 0,6 mM) que estão totalmente ocupados porque a concentração intra-íon (12 mM) permite.
Neste estado (E1) e com o Na + ligado aos seus sítios de ligação, o ATP se liga ao seu sítio no setor citosólico da molécula, hidrolisa e um grupo fosfato é transferido para o aspartato 376, formando um acilfosfato de alta energia que induz um processo conformacional mudar para o estado E2.
A mudança conformacional para o estado E2 implica que os sítios de ligação do Na + passam para o exterior, sua afinidade pelo íon diminui muito e é liberado no fluido extracelular, enquanto, ao mesmo tempo, a afinidade dos sítios de ligação do K + aumenta e esses íons são anexados à parte externa da bomba.


Uma vez que o Na + é liberado e o K + ligado, ocorre a hidrólise do fosfato de aspartilo e a mudança conformacional do estado E2 para o estado E1 é revertida, com a reintrodução dos sítios de ligação do Na + vazios e do K + ocupado.
Quando essa mudança ocorre, os sítios do Na + recuperam sua afinidade e os do K + a perdem, com o que o K + é liberado na célula..
A bomba Na + / K + está presente na maioria, senão em todas as células de mamíferos, nas quais é de importância geral por ajudar a manter sua osmolaridade e, portanto, seu volume..
A entrada contínua de íons sódio na célula determina um aumento no número intracelular de partículas osmoticamente ativas, o que induz a entrada de água e o aumento de volume que acabaria causando o rompimento da membrana e o colapso celular..
Como essas bombas introduzem apenas 2 K + para cada 3 Na + retirados, elas se comportam eletrogenicamente, o que significa que "descompensam" as cargas elétricas internas, favorecendo a produção do potencial de membrana característico das células do corpo..
Sua importância fica evidente também em relação às células que compõem os tecidos excitáveis, nos quais os potenciais de ação se caracterizam pela entrada do íon Na +, que despolariza a célula, e pela saída do K +, que a repolariza..
Esses movimentos iônicos são possíveis graças ao funcionamento das bombas Na + / K +, que contribuem para a produção dos gradientes químicos que movem os íons envolvidos..
Sem essas bombas, operando na direção oposta, os gradientes de concentração desses íons se dissipariam e a atividade excitatória desapareceria..
Outro aspecto que destaca a extrema importância das bombas de sódio-potássio está relacionado à função renal, que sem elas seria impossível.
A função renal envolve a filtração diária de mais ou menos 180 litros de plasma e grandes quantidades de substâncias, algumas das quais devem ser excretadas, mas muitas devem ser reabsorvidas para que não sejam perdidas pela urina..
A reabsorção de sódio, água e muitas das substâncias filtradas depende dessas bombas, que estão localizadas na membrana basolateral das células que compõem os epitélios dos diferentes segmentos tubulares dos néfrons renais..
As células epiteliais que revestem os túbulos renais têm uma face que está em contato com o lúmen do túbulo e é chamada de face apical, e outra que está em contato com o interstício ao redor do túbulo e chamada de basolateral..
A água e as substâncias que são reabsorvidas devem primeiro passar para a célula através do apical e, em seguida, para o interstício através do basolateral.
A reabsorção do Na + é fundamental tanto em relação a ele, como em relação à água e às demais substâncias que dela dependem. A entrada apical do Na + na célula exige que haja um gradiente que o mova e isso implica uma concentração muito baixa do íon dentro da célula..
Essa baixa concentração intracelular de Na + é produzida pelas bombas de sódio da membrana basolateral, que atuam intensamente para remover o íon das células para o interstício..



Ainda sem comentários