Quitridiomicote É um filo que agrupa fungos microscópicos com esporos flagelados. Eles são possivelmente o grupo ancestral dos fungos superiores. Sua taxonomia é complicada devido à plasticidade de suas formas em diferentes condições de cultivo..
Eles são nutridos pela absorção. Existem saprófitas e parasitas, alguns dos quais são patógenos em plantações como a batata e outros causam estragos nas populações de anfíbios. Eles vivem no solo e também em águas doces e salgadas. Algumas espécies anaeróbicas obrigatórias habitam o trato digestivo dos herbívoros.
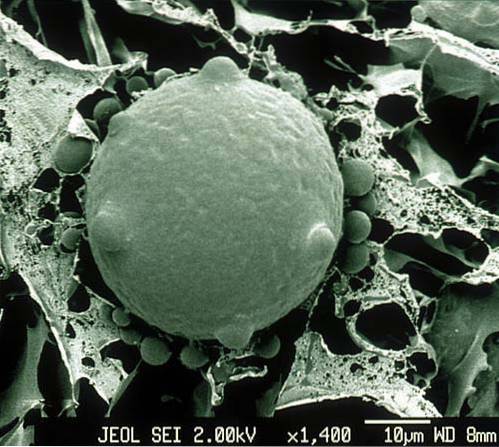
Eles se reproduzem assexuadamente por zoósporos, dando origem a uma fase haplóide na qual os gametas ciliados são produzidos. Então, por plasmogamia e cariogamia, é gerada uma fase diplóide que dá origem ao esporotal..
Índice do artigo
Eles são principalmente fungos filamentosos com hifas multinucleadas sem septos (cenocíticos). Embora também existam espécies unicelulares ou unicelulares com rizoides (raízes falsas com função de absorção formadas por filamentos curtos sem núcleo).
Suas paredes celulares são compostas de quitina, embora a celulose também exista em algumas espécies. Eles não formam colônias de hifas ramificadas (micélios) como acontece com os fungos não flagelados. Eles produzem corpos esferoidais multinucleados chamados talos.
Os talos são as estruturas de alimentação que mais tarde se transformam em esporrângios. O esporângio é uma estrutura semelhante a um saco de paredes finas, em cujo protoplasma são formados zoósporos..
Chytridiomicotas apresentam formas muito variadas de estrutura do talo e dos órgãos reprodutivos. Algumas espécies têm talo holocárpico (ele se transforma completamente em esporrangia).
Outras formas mostram talos eucarpicos (mantém funções vegetativas e reprodutivas) e podem ser monocêntricas (esporrângios com rizóides) ou policêntricos (vários esporrângios e rizóides). As formas mais simples são endoparasitas.
O mais relevante desses fungos é a produção de células móveis: esporos flagelados e gametas ciliados. Zoósporos são células sem parede celular, geralmente de 2 a 10 μm de diâmetro, que contêm um único núcleo..
Com exceção de alguns gêneros de Neocallimastigales, os zoósporos são impulsionados por um único flagelo liso orientado posteriormente..
O zoósporo fornece caracteres importantes para a classificação de fungos flagelados. Graças ao uso do microscópio eletrônico, várias estruturas peculiares foram detectadas. Entre eles estão: o rumposome e a tampa nuclear.
A liberação de esporos geralmente ocorre por meio de poros sem tampa (sem tampa) ou tubos de descarga e raramente através de aberturas tampadas.
Nos zoósporos, existe um conjunto de membranas cisternas fenestradas ou não fenestradas, denominado rumpossoma, cuja função é desconhecida. A capa nuclear é uma agregação de ribossomos ligados ao núcleo e cobertos por um prolongamento da membrana nuclear.
A quitridiomicose é uma doença que afeta anfíbios, principalmente sapos e rãs. É causado pelo fungo Batrachochytrium dendrobatidis, apenas Chytridiomicota conhecido por ser um parasita de vertebrados.
A pele dos animais fica infectada quando eles entram em contato com águas onde os zoósporos do fungo são encontrados. Em alguns casos, esse fungo exterminou 100% da população de rãs e chegou a ser a causa da extinção de quatro espécies de rãs na Austrália..
Pensa-se que as enzimas secretadas pelo fungo afetam a estrutura epidérmica impedindo as diversas funções desse tecido na rã (regulação osmótica, absorção de água, respiração), o que acaba sendo fatal na maioria dos casos.
Espécies de gênero Coelomyces Eles parasitam larvas de mosquitos, por isso são importantes para o controle biológico desses insetos.
Existem também várias espécies de quitridiomicos parasitas obrigatórios de uma grande variedade de plantas vasculares. Espécies de gênero Synchytrium eles atacam a cultura da batata.
Synchytrium endobioticum é um fungo endobiótico que produz a chamada “verruga da batata preta”, uma doença importante da cultura. O genero Olpidium inclui espécies que afetam várias cucurbitáceas.
Esta doença é causada quando os zoósporos penetram no tecido dos tubérculos, invadindo as células e utilizando-as como receptores reprodutivos..
O ciclo reprodutivo é repetido continuamente no tecido da batata, causando a proliferação celular. Os tubérculos adquirem aspecto verrucoso, semelhante à proliferação dos botões florais.
Devido ao seu tamanho microscópico, os quitridiomicotes não podem ser observados diretamente na natureza. Eles só podem ser detectados por análise microscópica de tecidos ou restos vegetais, pele de anfíbios afetados por algumas espécies desses fungos, água ou amostras de solo..
Os especialistas nesses fungos desenvolveram técnicas especiais para coletá-los e posteriormente cultivá-los em laboratório em meio artificial..
O filo Chytridiomicota é um grupo monofilético considerado uma das linhagens basais da árvore evolutiva do reino Fungi. Compreende uma única classe (Chytridiomicetos), tradicionalmente dividida em cinco ordens: Chytridiales, Spizellomycetales, Blastocladiales, Monoblepharidales e Neocallimastigales.
Os critérios para a separação dessas ordens são principalmente o modo de reprodução e a ultraestrutura do zoósporo. Inclui cerca de 1000 espécies.
Monoblofaridales são oogâmicos (gameta feminino imóvel fertilizado por um gameta masculino móvel). Os Blastocladiales apresentam meiose espórica e alternância de gerações esporofíticas e gametofíticas..
Chytridiales são caracterizados por sua meiose zigótica (indivíduos haplóides). Spizellomycetales são definidos por caracteres ultraestruturais peculiares.
Neocallimastigales incluem exclusivamente simbiontes anaeróbicos do rúmen de herbívoros e com zoósporos multiflagelados.
Recentemente, foi proposto separar Blastocladiales e Neocallimastigales deste grupo, elevando-os à categoria de filo. Enquanto Monoblepharidales é reduzido à categoria de classe.
Ao mesmo tempo, uma nova ordem de Chytridiomicota foi postulada: Lobulomycetales. Esta nova ordem é baseada principalmente na análise genética, em menor medida em dados ultraestruturais e morfologia..
Eles se alimentam absorvendo o substrato por meio de seus rizóides. Essas estruturas secretam enzimas que digerem o substrato e as substâncias migram em direção às hifas de absorção. Existem saprófitas e parasitas.
As quitridiomicotas são classificadas de acordo com a posição que ocupam em relação ao substrato: no substrato (epibióticos) ou dentro do substrato (endobióticos).
Eles vivem no solo em decomposição de matéria orgânica, na superfície de plantas ou animais e também na água. Eles estão localizados desde áreas tropicais até o Ártico.
Algumas espécies são parasitas de outros fungos, como algas, plâncton e plantas vasculares, além de animais.
Na água doce, eles são encontrados em riachos, lagoas e estuários. Em ecossistemas marinhos, principalmente como parasitas de algas e componentes do plâncton. As espécies de quitridiomicote estão localizadas desde sedimentos costeiros até profundidades do mar de 10.000 m.
Talvez a maioria das espécies de Chytridiomicota sejam encontradas em habitats terrestres como florestas, solos agrícolas e desérticos e em pântanos ácidos como saprotróficos de substratos refratários como pólen, quitina, queratina e celulose no solo..
Algumas espécies habitam o interior dos tecidos vegetais como parasitas obrigatórios; tanto nas folhas, caules e raízes.
Espécies da ordem Neocallimastigales vivem no rúmen de animais herbívoros. Devido à sua capacidade de degradar a celulose, eles desempenham um papel importante no metabolismo ruminal..
O ciclo de vida dos quitridiomicos tem uma fase assexuada e outra sexual.
Na fase assexuada, a reprodução pode ser planogamética. A partir de um esporângio de resistência ou latência, são produzidos zoósporos, ou seja, esporos flagelados móveis haplóides..
Os zoósporos nadam livremente por algum tempo e se transformam em cistos que, ao germinar em condições favoráveis, dão origem a um gametotalus..
Aqui se formam os gametângios masculino e feminino que darão origem aos respectivos planogametas, iniciando a fase sexual. Esses gametas possuidores de cílios são móveis.
Na fase sexual, por fusão de citoplasmas (plasmogamia) e posterior fusão dos núcleos (cariogamia) dos planogametas, forma-se o zigoto. É aqui que começa a fase diplóide do ciclo.
O zigoto pode assumir a forma de um cisto de resistência ou se transformar em um talo cenocítico diplóide (esporotal).
Os esporotais dão origem a esporrângios de resistência que formam zoósporos haplóides, fechando assim o ciclo. A partir do esporotélio, também podem se formar zoosporângios que produzem zoósporos diplóides que encistam e, ao germinar, formam novos esporotélios..
A reprodução sexual também pode ocorrer por oogamia, como é o caso na ordem Monoblefaridales. Aqui, é um gameta feminino imóvel localizado no oogônio que recebe o gameta masculino flagelado móvel..
Os quitriomicotos parasitas geralmente têm um ciclo mais simples.
Em fungos parasitas de plantas, como Olpidium viciae, o zigoto é móvel e infecta a planta hospedeira. Os esporrângios de resistência desenvolvem-se dentro das células vegetais. Dentro desses esporrângios ocorre cariogamia.
A germinação finalmente ocorre e os zoósporos são liberados. Os zoósporos também podem infectar diretamente a planta hospedeira.
No cogumelo Batrachochytrium dendrobatidis parasita da pele de rã, após um período de mobilidade inferior a 24 horas os zoósporos reabsorvem seus flagelos e encistos. Eles então germinam formando rizóides que aderem à pele da rã.
O talo se desenvolve em um esporângio que dá origem a uma nova geração de zoósporos. A fase sexual ainda é desconhecida.
Ainda sem comentários