O cromatina É o complexo formado por DNA e proteínas, único em organismos eucarióticos. Em termos de proporção, contém quase duas vezes mais proteínas do que material genético. As proteínas mais importantes neste complexo são as histonas - pequenas proteínas carregadas positivamente que se ligam ao DNA por meio de interações eletrostáticas. Além disso, a cromatina tem mais de mil proteínas além das histonas..
A unidade fundamental da cromatina é o nucleossomo, que consiste na união de histonas e DNA. Este arranjo é uma reminiscência das contas de um colar. Depois de passar por todos os níveis superiores de organização do DNA, chegaremos aos cromossomos.
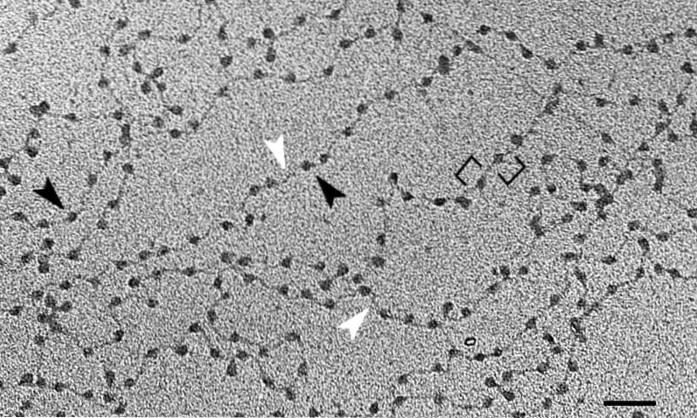
A estrutura da cromatina está intimamente relacionada ao controle da expressão gênica. Existem dois tipos principais: eucromatina e heterocromatina.
A eucromatina é caracterizada por ter um baixo grau de compactação, o que se traduz em altos níveis de transcrição. Em contraste, a heterocromatina é transcricionalmente inativa, devido ao seu alto grau de compactação..
Estruturalmente, existem certas marcas epigenéticas das histonas típicas de ambos os tipos de cromatinas. Enquanto a eucromatina está associada à acetilação, a heterocromatina está associada a uma diminuição nos grupos químicos.
Existem algumas regiões de heterocromatina com funções estruturais, como telômeros e centrômeros.
Índice do artigo
O estudo do material genético e sua organização estrutural teve início em 1831, quando o pesquisador Robert Brown descreveu o núcleo. Uma das questões imediatas a essa descoberta era indagar sobre a natureza biológica e química dessa estrutura..
Essas questões começaram a ser elucidadas entre 1870 e 1900, com os experimentos de Friedrich Miescher, que introduziu a palavra nucleína. No entanto, Walther Flemming modifica o termo e usa cromatina para se referir à substância nuclear.
Com o passar do tempo, você começa a ter uma compreensão mais profunda do material genético e de suas propriedades. Só em 1908 o pesquisador italiano Pasquale Baccarini percebeu que a cromatina não era homogênea e conseguiu visualizar pequenos corpos dentro do núcleo..
Os tipos de cromatinas - eucromatina e heterocromatina - foram inicialmente propostos por Emil Heitz em 1928. Para estabelecer essa classificação, Heitz baseou-se no uso de corantes.
Em 1974, o biólogo Roger Kornberg propôs um modelo de organização do material genético em estruturas conhecidas como nucleossomos, hipótese confirmada empiricamente pelos experimentos de Markus Noll..
A cromatina é uma nucleoproteína formada pela união de material genético - DNA - com um conjunto heterogêneo de proteínas. Esta associação é altamente dinâmica e adquire uma conformação tridimensional complexa que lhe permite cumprir as suas funções reguladoras e estruturais..
Uma das proteínas da cromatina mais importantes são as histonas, que são encontradas quase na mesma proporção do DNA..
Histonas são proteínas básicas, notavelmente conservadas ao longo da história evolutiva dos seres orgânicos - ou seja, nossas histonas não variam muito em comparação com as de outros mamíferos, mesmo outro animal mais distante filogeneticamente.
A carga das histonas é positiva, portanto, elas podem interagir por meio de forças eletrostáticas com a carga negativa do esqueleto de fosfato presente no DNA. Existem cinco tipos de histonas, a saber: H1, H2A, H2B, H3 e H4.
Existem também várias proteínas de natureza diferente das histonas que participam da compactação do DNA..
A unidade básica da cromatina são os nucleossomos - estruturas repetitivas compostas de DNA e histonas, uma conformação encontrada em todo o material genético..
A dupla hélice do DNA é enrolada em um complexo de oito histonas conhecido como histona octâmero. A molécula se enrola em aproximadamente duas voltas, seguida por uma região curta (entre 20 e 60 pares de bases) que separa os nucleossomos uns dos outros..
Para entender essa organização, devemos levar em conta que a molécula de DNA é extremamente longa (cerca de 2 metros) e deve ser enrolada de forma ordenada para se estabelecer no núcleo (cujo diâmetro é de 3 a 10 µm). Além disso, deve ser disponibilizado para replicação e transcrição..
Este objetivo é alcançado com diferentes níveis de compactação do DNA, sendo o primeiro os supracitados nucleossomos. Assemelham-se às contas de um colar de pérolas. Aproximadamente 150 pares de bases de DNA são "contados" em casa.
Não existem histórias verdadeiras sobre bactérias. Em contraste, há uma série de proteínas que se assemelham às histonas e presume-se que contribuam para o empacotamento do DNA bacteriano..
A organização da cromatina não é restrita ao nível do nucleossomo. Essa associação de proteínas e DNA são agrupadas em uma estrutura mais espessa de cerca de 30 nm - devido a essa espessura é chamada de nível de "fibra de 30 nm"..
A cromatina organizada na espessura de 30 nm é organizada por sua vez na forma de loops que se estendem em uma espécie de arcabouço de natureza proteica (não histonas)..
Este é o modelo atualmente em uso, embora seja previsível a existência de mecanismos de compactação mais complexos. A organização final consiste no cromossomo.
A compactação e organização do material genético são vitais para múltiplas funções biológicas. Várias condições médicas foram associadas a erros na estrutura da cromatina, incluindo talassemia alfa ligada ao X, síndrome de Rubinstein-Taybi, síndrome de Coffin-Lowry, síndrome de Rett, entre outras..
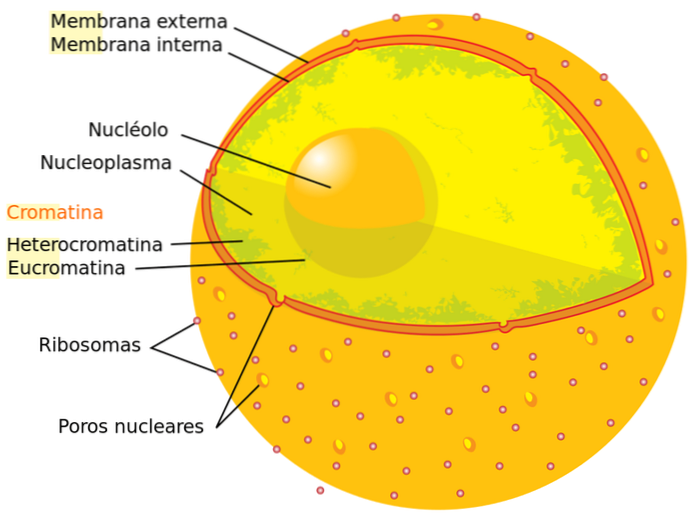
Existem dois tipos de cromatina na célula, revelados pela aplicação de corantes: eucromatina (cromatina "verdadeira") e heterocromatina. No primeiro caso, a coloração é fracamente observada, enquanto no segundo a coloração é intensa.
Essa organização estrutural do DNA é exclusiva dos organismos eucarióticos e é crucial para o comportamento dos cromossomos e a regulação da expressão gênica..
Se avaliarmos as proporções de ambos os tipos de cromatinas em uma célula que está em interfase, descobrimos que aproximadamente 90% da cromatina é eucromatina e os 10% restantes correspondem à heterocromatina. Descreveremos cada tipo em detalhes abaixo:
A principal diferença que existe entre os dois tipos de cromatina está relacionada ao grau de compactação ou "empacotamento" da molécula durante estágios específicos da divisão celular..
Embora na interface o material genético pareça estar disperso aleatoriamente, não é desta forma.
Há uma organização significativa neste estágio, onde uma partição diferencial do material cromossômico pode ser vista dentro do núcleo.
O DNA dos cromossomos não se entrelaça com a fita de DNA de outros cromossomos e permanece em regiões específicas chamadas territórios cromossômicos. Esta organização parece contribuir para a expressão gênica.
A heterocromatina é fortemente condensada, tornando-a inacessível ao mecanismo de transcrição - portanto, não é transcrita. Além disso, é pobre em termos de número de genes..
Certas regiões de heterocromatina são persistentes ao longo das linhagens celulares - ou seja,, para sempre ele se comportará como heterocromatina. Esse tipo de heterocromatina é conhecido como constitutivo. Um exemplo disso são as regiões condensadas dos cromossomos chamados centrômeros e telômeros..
Em contraste, existem porções de heterocromatina que podem variar o nível de compactação em resposta a mudanças nos padrões de desenvolvimento ou variáveis ambientais..
Graças a novos estudos, essa visão está sendo reformulada e agora há evidências de que a heterocromatina constitutiva também é dinâmica e capaz de responder a estímulos..
Um dos fatores que determinam a estrutura da cromatina são as modificações químicas das histonas. No caso da cromatina que é transcricionalmente inativa, eles exibem histonas hipoacetiladas.
A diminuição do número de grupos acetila está associada ao silenciamento gênico, uma vez que não mascara a carga positiva das lisinas, permitindo uma forte interação eletrostática entre o DNA e as histonas..
Outra marca epigenética é a metilação. Porém, como a adição de um grupo metil não modifica a carga da proteína, sua consequência (genes ativadores ou desativadores) não é tão óbvia e dependerá da região da histona onde a marca se encontra..
Empiricamente, descobriu-se que a metilação de H3K4me3 e H3K36me3 está associada à ativação do gene e de H3K9me3 e H3K27me3.
No exemplo da heterocromatina constitutiva mencionamos o centrômero. Esta região cromossômica tem um papel estrutural e contribui para o movimento dos cromossomos durante os eventos de divisão celular mitótica e meiótica..
Ao contrário da heterocromatina, a eucromatina é uma molécula menos compacta, portanto, a maquinaria de transcrição tem fácil acesso (especificamente a enzima RNA polimerase) e pode ser ativamente expressa por meio de vias genéticas.
A estrutura do nucleossomo de uma cromatina transcricionalmente ativa é caracterizada por histonas acetiladas. A presença de lisinas monometiladas também está associada à ativação do gene.
A adição de um grupo acetil a esses resíduos de histona lisina neutraliza a carga positiva do referido aminoácido. A consequência imediata dessa mudança é a redução das interações eletrostáticas entre histonas e DNA, gerando uma cromatina mais frouxa..
Essa modificação estrutural permite as interações do material genético com a maquinaria transcricional, que se caracteriza por ser particularmente volumosa..
A eucromatina engloba todos os genes que estão ativos e conseguem ser alcançados pela maquinaria enzimática relacionada à transcrição. Portanto, as funções são tão amplas quanto as funções dos genes envolvidos..



Ainda sem comentários