O eritropoiese É o processo pelo qual os glóbulos vermelhos ou eritrócitos são formados. Essas células sanguíneas, em humanos, têm vida média de 4 meses e são incapazes de se reproduzir. Devido a isso, novos eritrócitos devem ser criados para substituir aqueles que morrem ou se perdem nas hemorragias..
Nos homens, o número de glóbulos vermelhos é de aproximadamente 54 milhões por mililitro, enquanto nas mulheres é um pouco menor (48 milhões). Cerca de 10 milhões de eritrócitos são perdidos diariamente, portanto, uma quantidade semelhante deve ser reposta.
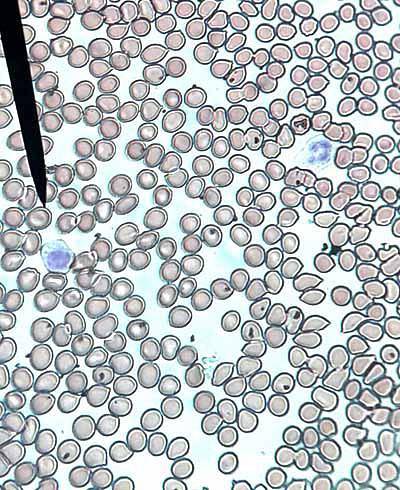
Os eritrócitos são formados a partir de eritroblastos nucleados presentes na medula óssea vermelha de mamíferos, enquanto em outros vertebrados são produzidos principalmente nos rins e no baço..
Quando chegam ao fim de seus dias, eles se fragmentam; então, as células chamadas macrófagos os englobam. Esses macrófagos estão presentes no fígado, medula óssea vermelha e baço..
Quando os glóbulos vermelhos são destruídos, o ferro é reciclado para ser usado novamente, enquanto o resto da hemoglobina é transformada em um pigmento biliar chamado bilirrubina..
A eritropoiese é estimulada por um hormônio denominado eritropoietina, mas o processo é regulado por diversos fatores, como temperatura, pressão de oxigênio, entre outros..
Índice do artigo
Em organismos adultos, a eritropoiese ocorre em locais especializados na medula óssea vermelha chamados ilhas eritroblásticas. Para a formação dos eritrócitos, vários processos devem ocorrer, desde a proliferação celular até a maturação das hemácias, passando por vários estágios de diferenciação celular..
À medida que as células sofrem divisões mitóticas, seu tamanho e o do núcleo diminuem, bem como a condensação da cromatina e a hemoglobinização. Além disso, eles estão se afastando da área de origem.
Nos estágios finais, perderão o núcleo e outras organelas e entrarão na circulação, migrando pelos poros citoplasmáticos das células endoteliais..
Alguns autores dividem todo o processo de eritropoiese em duas fases, a primeira de proliferação e diferenciação celular; enquanto outros dividem o processo com base nas características específicas da célula em cada estágio, quando observada com a coloração de Wright. Com base neste último, os estágios da eritropoiese são:
São as primeiras células sensíveis à eritropoietina, alguns autores as chamam de progenitoras mieloides, ou também BFU-E, por sua sigla em inglês. Eles são caracterizados por expressar antígenos de superfície como o CD34, bem como pela presença de receptores de eritropoietina em pequenas quantidades..
Abreviado em inglês como CFU-E, eles são capazes de produzir pequenas colônias de eritroblastos. Outra característica dessas células é que as quantidades de receptores de eritropoietina são muito maiores do que em unidades formadoras de colônias estouradas..
Considerado o primeiro estágio de maturação dos eritrócitos. São caracterizados por seu grande tamanho (14 a 19 µm segundo alguns autores, até 25 µm segundo outros). O núcleo é arredondado e também apresenta nucléolos e cromatina abundante.
Considerado o primeiro estágio de maturação dos eritrócitos. São caracterizados por seu grande tamanho (14 a 19 µm segundo alguns autores, até 25 µm segundo outros). O núcleo é grande, arredondado, com cromatina disposta em forma de filamentos e 2 ou 3 nucléolos.
A captação de ferro no plasma começa nesta fase. Eles têm meia-vida de 20 horas, para passar pela mitose ao próximo estágio.
Também chamados de normoblastos, eles são menores que seus precursores. Essas células se coram de azul com a coloração vital, ou seja, são basofílicas. O núcleo está condensado, os nucléolos desapareceram e eles têm um grande número de ribossomos. Nesta fase, a síntese da hemoglobina começa.
No início são conhecidos como eritroblastos basofílicos Tipo I e após uma divisão mitótica transformam-se no Tipo II, que permanecem basófilos e apresentam maior síntese de hemoglobina. A duração aproximada de ambas as células, juntas, é semelhante à dos proeritroblastos.
São formadas por divisão mitótica de eritroblastos basofílicos do tipo II e são as últimas células capazes de se dividir por mitose. Seu tamanho varia de 8 a 12 µm, e eles têm um núcleo arredondado e condensado.
O citoplasma dessas células se cora em cinza chumbo com a coloração de Wright. Possui alta concentração de hemoglobina e o número de ribossomos permanece alto.
6-eritroblastos ortocromáticos
A cor dessas células é rosa ou vermelha devido à quantidade de hemoglobina que possuem. Seu tamanho é um pouco menor que o de seus precursores (7 a 10 µm) e possui um núcleo pequeno, que será expelido por exocitose quando as células amadurecerem..
São formados pela diferenciação de eritroblastos ortocrômicos, perdem organelas e preenchem seu citoplasma com hemoglobina. Permanecem na medula óssea vermelha por dois a três dias até que migrem para o sangue, onde completam sua maturação..
Eles são os elementos formados maduros, o produto final da eritropoiese e que são formados pela maturação dos reticulócitos. Possuem formato bicôncavo devido à ausência de núcleo e à interação entre o citoesqueleto eritrocitário e duas proteínas denominadas espectrina e actina..
São as células sanguíneas mais abundantes, formadas a partir de reticulócitos. Em mamíferos, apresentam formato bicôncavo devido à ausência de núcleo e à interação entre o citoesqueleto eritrocitário e duas proteínas chamadas espectrina e actina. Em outros vertebrados, eles são arredondados e retêm o núcleo.
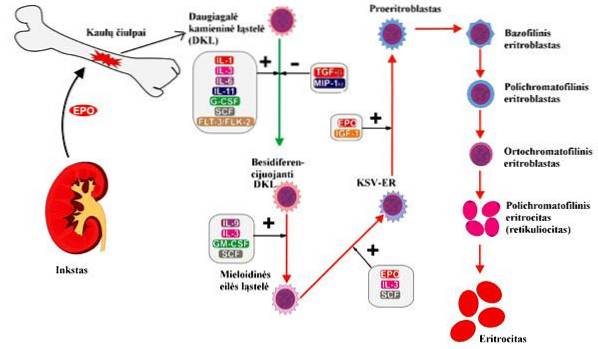
Embora a eritropoietina estimule a formação de glóbulos vermelhos para melhorar a capacidade de transporte de oxigênio do sangue, existem vários mecanismos fundamentais para regular essa formação, incluindo:
A concentração de oxigênio no sangue regula a eritropoiese. Quando essa concentração é muito baixa no fluxo sanguíneo para o rim, a produção de glóbulos vermelhos é estimulada..
Essa baixa concentração de O2 tecidual pode ocorrer devido à hipoxemia, anemia, isquemia renal ou quando a afinidade da hemoglobina por esse gás é maior que o normal..
Miescher, em 1893, foi o primeiro a sugerir a relação entre hipóxia tecidual e eritropoiese. No entanto, essa hipóxia não estimula diretamente a medula óssea a produzir glóbulos vermelhos, como sugeriu Miescher. Pelo contrário, induz o rim a produzir o hormônio eritropoietina.
A produção de eritropoietina devido à hipóxia tecidual é regulada geneticamente, e os receptores que detectam essa hipóxia são encontrados no rim. A produção de eritropoietina também aumenta devido a uma queda na pressão parcial de oxigênio nos tecidos após o sangramento..
As células que produzem eritropoietina são encontradas nos rins e no fígado. O aumento da produção desse hormônio durante a anemia se deve ao aumento do número de células que o produzem..
A testosterona regula indiretamente a eritropoiese, regulando os níveis de ferro no sangue. Esse hormônio atua diretamente sobre a ação de uma proteína citoplasmática chamada BMP-Smad (proteína morfogenética óssea-Smad) nos hepatócitos.
Devido à ação da testosterona, a transcrição da hepcidina é suprimida. Esta hepcidina impede a passagem de ferro das células para o plasma de macrófagos que reciclam o ferro, levando a uma diminuição drástica do ferro no sangue.
Quando ocorrer hipoferremia, haverá inibição da eritropoietina, pois não haverá ferro para a produção de eritrócitos..
A temperatura demonstrou ter um efeito sobre a eritripoiese. As exposições a temperaturas muito baixas causam a necessidade de produzir calor nos tecidos.
Isso requer o aumento do número de eritrócitos para fornecer oxigênio aos tecidos periféricos. No entanto, não está totalmente claro como esse tipo de regulação ocorre..
Aparentemente, há produção de eritropoietina pelos neurônios do sistema nervoso central, para se protegerem do dano isquêmico e da apoptose. No entanto, os cientistas ainda não foram capazes de verificar isso..
Os agentes estimuladores da eritropoiese (AEEs) são agentes responsáveis por estimular a produção de eritrócitos. A eritropoietina é o hormônio naturalmente responsável por esse processo, mas também existem produtos sintéticos com propriedades semelhantes..
A eritropoietina é um hormônio sintetizado principalmente no rim. Durante os primeiros estágios de desenvolvimento, o fígado também está envolvido na produção ativa de eritropoietina. No entanto, à medida que o desenvolvimento avança, o último órgão tem menos envolvimento no processo..
O eritrócito começa a dispersar receptores para eritropoietina na superfície da membrana. A eritropoietina ativa uma série de cascatas de transdução de sinal intercelular que inicialmente produzem a síntese de hemoglobina e fazem com que os reticulócitos atuem mais rapidamente e sejam liberados na circulação..
Os AEE artificiais são classificados em gerações (da primeira à terceira), dependendo da data em que foram criados e comercializados. Estrutural e funcionalmente semelhante à eritropoietina.
Os ESAs de primeira geração são conhecidos como epoetina alfa, beta e delta. Os dois primeiros são produzidos por recombinação de células animais e têm meia-vida de cerca de 8 horas no corpo. A epetina delta, por sua vez, é sintetizada a partir de células humanas.
A darbepoetina alfa é um AEE de segunda geração, produzido a partir de células de hamster chinês usando tecnologia chamada DNA recombinante. Tem meia-vida três vezes mais longa do que os ESAs de primeira geração. Tal como acontece com as epoetinas, alguns atletas de alto desempenho têm usado a darbepoetina como meio de doping.
Continuous Erythropoetin Receptor Activator, ou CERA por sua sigla em inglês, é o nome genérico para ESAs de terceira geração. Eles não tentam simular a estrutura e função da eritropoietina, mas agem estimulando o receptor da eritropoietina, aumentando assim seus efeitos..
Sua meia-vida é de várias semanas em vez de horas, como os medicamentos anteriores. Usado comercialmente desde 2008, no entanto, seu uso ilícito em atividades esportivas aparentemente data de dois ou três anos antes de sua comercialização legal.
A eritropoiese ineficaz ou ineficaz ocorre quando os glóbulos vermelhos formados são defeituosos e geralmente são destruídos antes ou logo após deixarem a medula óssea..
A eritropoiese ineficaz pode ser decorrente de defeitos na síntese de ácidos nucléicos, do grupo heme ou das globinas. Esses defeitos causam diferentes tipos de anemia.
Nesse caso, há deficiência de ácido fólico e cobalamina, a síntese de DNA é inibida no núcleo das células promotoras dos eritrócitos, de modo que não conseguem se dividir mitoticamente. O citoplasma, por sua vez, aumenta seu volume (macrocitose), originando uma grande célula denominada megaloblasto..
Nestes casos, origina-se uma série de anemias denominadas anemias megaloblásticas, das quais a mais comum é a anemia perniciosa. Nesta doença, não há absorção de vitamina B12 no intestino delgado.
Outras causas de anemia megaloblástica incluem doenças digestivas, má absorção, deficiência de ácido fólico e devido a certos medicamentos.
Os sintomas desse tipo de anemia incluem palidez anormal, irritabilidade, perda de apetite, diarreia, dificuldade para andar ou fraqueza muscular. Dependendo da causa, pode ser tratada com suplementos vitamínicos ou de ácido fólico..
A eritropoiese ineficaz devido à deficiência na síntese de ferro pode causar dois tipos de anemia; anemia microcítica por deficiência de ferro e anemia sideroblástica.
A anemia microcítica é conhecida como um grupo de anemias caracterizadas por glóbulos vermelhos pequenos e claros, podendo ter diferentes origens, incluindo talassemia e eritropoiese ineficaz.
Os níveis de ferro e hemossiderina são muito altos na anemia sideroblástica. A hemossiderina é um pigmento amarelo derivado da hemoglobina e aparece quando os níveis do metal estão acima do normal. Este tipo de anemia causa a morte de basófilos da medula óssea vermelha e não há síntese de hemoglobina.
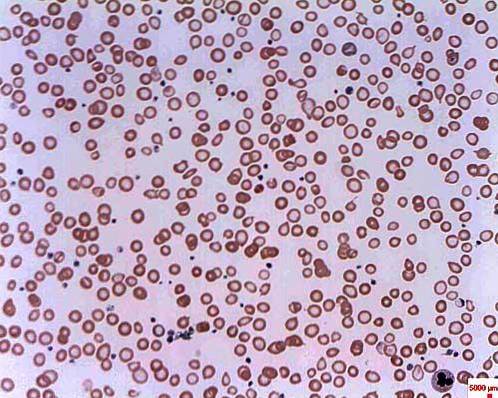
É chamada de anemia sideroblástica porque os eritrócitos se desenvolvem de forma anormal devido ao acúmulo de ferro na forma de grânulos, recebendo o nome de sideroblastos. A anemia sideroblástica pode ser congênita ou secundária e ter diferentes causas.
Nesse caso, ocorrem anemia falciforme e beta talassemia. A anemia falciforme também é conhecida como anemia falciforme. É produzida por uma mutação genética que leva à substituição do ácido glutâmico pela valina durante a síntese da beta globina..
Com essa substituição, diminui a afinidade da hemoglobina pelo oxigênio e ocorre atrofia do eritrócito, adquirindo formato de foice ao invés do formato de disco bicôncavo normal. O paciente com anemia falciforme é suscetível a microinfartos e hemólise.
A talassemia é uma doença causada por uma codificação genética inadequada de α e β-globinas que leva à morte precoce do eritrócito. Existem cerca de uma centena de mutações diferentes que podem causar talassemia com diferentes graus de gravidade.


Ainda sem comentários