UMA homozigoto na genética é um indivíduo que possui duas cópias do mesmo alelo (a mesma versão de um gene) em um ou mais loci (coloque no cromossomo). O termo é algumas vezes aplicado a entidades genéticas maiores, como cromossomos completos; neste contexto, um homozigoto é um indivíduo com duas cópias idênticas do mesmo cromossomo.
A palavra homozigota é composta de dois elementos etimologicamente. Os termos são homo -igual ou idêntico - e zigoto -óvulo fertilizado ou a primeira célula de um indivíduo originado por reprodução sexual-.
Índice do artigo
Os organismos são classificados com base em várias propriedades associadas ao material genético (DNA) contido em suas células. Considerando a estrutura celular onde o material genético está localizado, os organismos foram classificados em dois tipos principais: procariotos (pró: antes; karyon: núcleo) e eucariotos (eu: verdadeiro; karyon: testemunho).
Em organismos procarióticos, o material genético está confinado a uma região particular no citoplasma das células chamada nucleóide. Os organismos modelo neste grupo correspondem a bactérias da espécie Escherichia coli, que têm uma única fita circular de DNA, ou seja, suas extremidades são unidas.
Esta cadeia é conhecida como um cromossomo e em E. coli ele contém cerca de 1,3 milhão de pares de bases. Existem algumas exceções a este padrão dentro do grupo, por exemplo, alguns gêneros bacterianos têm cromossomos de cadeia linear, como espiroquetas do gênero Borrelia.
O tamanho ou comprimento linear dos genomas / cromossomos bacterianos está geralmente na faixa dos milímetros, ou seja, eles são várias vezes maiores do que o tamanho das próprias células..
O material genético é armazenado em uma forma embalada para reduzir o espaço ocupado por essa grande molécula. Esse empacotamento é obtido por meio do superenrolamento, uma torção no eixo principal da molécula que produz pequenas torções que, ao girar, causam.
Por sua vez, torções maiores desses pequenos fios em si mesmos e no resto da cadeia, reduzindo assim a distância e o espaço ocupado entre as diferentes seções do cromossomo circular e levando-o a uma forma condensada (dobrada).
Nos eucariotos, o material genético está localizado dentro de um compartimento especializado rodeado por uma membrana; Este compartimento é conhecido como núcleo da célula..
O material genético contido no núcleo é estruturado sob um princípio semelhante ao dos procariontes, o superenrolamento.
No entanto, os graus / níveis de torção são maiores, uma vez que a quantidade de DNA a ser acomodada é muito maior. Nos eucariotos, o núcleo não contém uma única cadeia de DNA ou cromossomo, ele contém vários deles e estes não são circulares, mas lineares e devem ser arranjados.
Cada cromossomo varia em tamanho dependendo da espécie, mas geralmente são maiores do que os procariontes quando comparados individualmente..
Por exemplo, o cromossomo humano 1 tem 7,3 centímetros de comprimento, enquanto o cromossomo de E. coli mede aproximadamente 1,6 milímetros. Para referência futura, o genoma humano contém 6,6 × 109 nucleotídeos.
Existe outra classificação de organismos baseada na quantidade de material genético que contêm, conhecida como ploidia..
Organismos com um único conjunto ou cópia de cromossomos são conhecidos como haplóides (bactérias ou células reprodutivas em humanos), com dois conjuntos / cópias de cromossomos são conhecidos como diplóides (Homo sapiens, Mus musculus, entre muitos outros), com quatro conjuntos / cópias de cromossomos são conhecidos como tetraplóides (Odontophrinus americanus, plantas do gênero Brassicca).
Organismos com grande número de conjuntos de cromossomos são conhecidos coletivamente como poliploides. Em muitos casos, os conjuntos extras de cromossomos são cópias de um conjunto básico.
Por vários anos, considerou-se que características como ploidia maior que um eram típicas de organismos com um núcleo celular definido, mas descobertas recentes mostraram que alguns procariotos têm múltiplas cópias cromossômicas aumentando sua ploidia, como mostrado pelos casos de Deinococcus radiodurans Y Bacillus meagateriium.
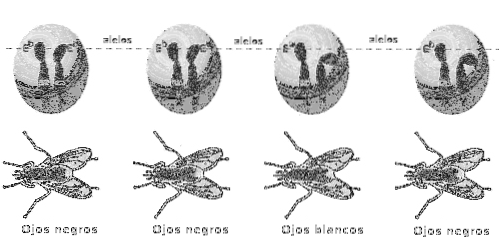
Em organismos diplóides (como as ervilhas estudadas por Mendel), os dois genes de um loci, ou alelos, um é herdado pela mãe e o outro pelo pai e o par de alelos juntos representa o genótipo desse gene específico.
Um indivíduo que apresenta um genótipo homozigoto (homozigoto) para um gene é aquele que possui duas variantes ou alelos idênticos em um locus dados.
Os homozigotos, por sua vez, podem ser subclassificados em dois tipos com base em sua relação e contribuição para o fenótipo: dominante e recessivo. Deve-se notar que ambas as expressões são propriedades fenotípicas.
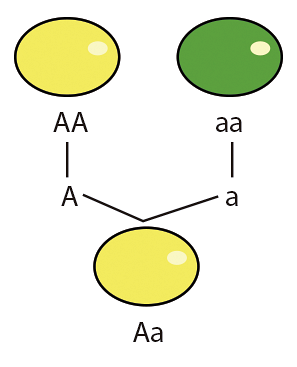
A dominância no contexto genético é uma relação entre os alelos de um gene em que a contribuição fenotípica de um alelo é mascarada pela contribuição do outro alelo do mesmo. locus; neste caso, o primeiro alelo é recessivo e o segundo é dominante (heterozigosidade).
A dominância não é herdada nos alelos ou no fenótipo que eles produzem, é uma relação que se estabelece a partir dos alelos presentes e pode ser modificada por agentes externos, como outros alelos..
Um exemplo clássico de dominância e sua relação com o fenótipo é a produção de uma proteína funcional pelo alelo dominante que finalmente produz o traço físico, enquanto o alelo recessivo não produz a referida proteína de forma funcional (mutante) e, portanto, não contribui para o fenótipo.
Assim, um indivíduo homozigoto dominante para um traço / característica é aquele que possui um genótipo que apresenta duas cópias idênticas do alelo dominante (linha pura).
Também é possível encontrar dominância em genótipos onde os dois alelos dominantes não são encontrados, mas um alelo dominante está presente e outro é recessivo, mas este não é um caso de homozigose, é um caso de heterozigosidade..
Na análise genética, os alelos dominantes são representados por uma letra maiúscula relacionada à característica que está sendo descrita..
No caso das pétalas da flor da ervilha, o traço selvagem (neste caso a cor roxa) é dominante e o genótipo é representado como "P / P", denotando tanto o traço dominante quanto a condição homozigótica, ou seja,, a presença de dois alelos idênticos em um organismo diplóide.
Por outro lado, um indivíduo homozigoto recessivo para um traço particular carrega duas cópias do alelo que codifica o traço recessivo.
Continuando com o exemplo da ervilha, o traço recessivo nas pétalas é a cor branca, então em indivíduos com flores desta cor cada alelo é representado com uma letra minúscula que implica recessividade e as duas cópias recessivas idênticas, então o genótipo é simbolizado como "p / p".
Em alguns casos, os geneticistas usam uma letra maiúscula simbolicamente para representar o alelo selvagem (por exemplo, P) e, assim, simbolizar e se referir a uma sequência de nucleotídeos específica.
Por outro lado, quando uma letra minúscula, p, é usada, ela representa um alelo recessivo que pode ser qualquer um dos tipos possíveis (mutações) [1,4,9].
Os processos pelos quais um determinado genótipo é capaz de produzir um fenótipo em organismos são variados e complexos. Mutações recessivas geralmente inativam o gene afetado e levam a uma perda de função.
Isso pode acontecer pela remoção parcial ou total do gene, pela interrupção da expressão do gene ou pela alteração da estrutura da proteína codificada que finalmente altera sua função..
Por outro lado, as mutações dominantes frequentemente produzem um ganho de função, podem aumentar a atividade de um determinado produto gênico ou conferir uma nova atividade ao referido produto e, portanto, também podem produzir expressão espaço-temporal inadequada..
Esse tipo de mutação também pode estar associado à perda de função, existem alguns casos em que duas cópias de um gene são necessárias para a função normal, de modo que a remoção de uma única cópia pode levar a um fenótipo mutante..
Esses genes são conhecidos como haplo-insuficiente. Em alguns outros casos, a mutação pode levar a mudanças estruturais nas proteínas que interferem com a função da proteína de tipo selvagem codificada pelo outro alelo. Estas são conhecidas como mutações negativas dominantes. .
Em humanos, exemplos de fenótipos recessivos conhecidos são albinismo, fibrose cística e fenilcetonúria. Todas essas são condições médicas com bases genéticas semelhantes..
Tomando este último como exemplo, os indivíduos com esta doença têm um genótipo “p / p” e, como o indivíduo possui ambos os alelos recessivos, é homozigoto.
Neste caso, o "p" está relacionado ao termo em inglês fenilcetonúria y é minúsculo para representar o caráter recessivo do alelo. A doença é causada pelo processamento anormal da fenilalanina, que em condições normais deve ser convertida em tirosina (ambas as moléculas são aminoácidos) pela enzima fenilalanina hidroxilase..
Mutações na proximidade do sítio ativo desta enzima impedem que ela seja capaz de se ligar à fenilalanina para processá-la posteriormente.
Como consequência, a fenilalanina se acumula no corpo e é convertida em ácido fenilpirúvico, um composto que interfere no desenvolvimento do sistema nervoso. Essas condições são conhecidas coletivamente como distúrbios autossômicos recessivos..
Os padrões de herança e, portanto, a presença de alelos para um gene, tanto dominante quanto recessivo, nos genótipos de indivíduos de uma população obedecem à primeira lei de Mendel.
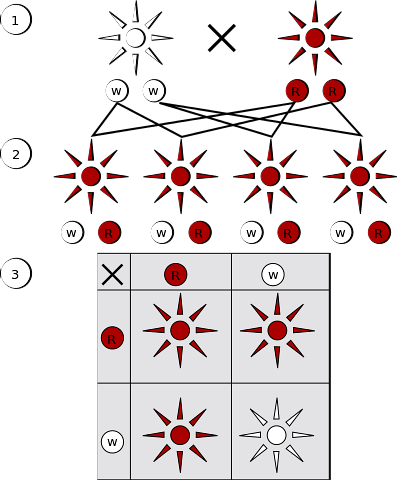
Essa lei é conhecida como lei da segregação igualitária de alelos e tem bases moleculares que são explicadas durante a formação dos gametas..
Em organismos diplóides que se reproduzem sexualmente, existem dois tipos principais de células: células somáticas e células sexuais ou gametas..
As células somáticas têm duas cópias de cada cromossomo (diplóide) e cada um dos cromossomos (cromátides) contém um dos dois alelos.
As células gaméticas são produzidas pelos tecidos germinativos através da meiose onde as células diplóides sofrem divisão nuclear acompanhada por uma redução cromossômica durante este processo, consequentemente elas possuem apenas um conjunto de cromossomos, portanto são haplóides..
Durante a meiose, o fuso acromático é ancorado aos centrômeros dos cromossomos e as cromátides são separadas (e, portanto, os alelos também) em pólos opostos da célula-tronco, produzindo duas células-filhas ou gametas separadas..
Se o indivíduo que produz os gametas for homozigoto (A / A ou a / a), o número total de células gaméticas produzidas por ele carregará alelos idênticos (A ou a, respectivamente).
Se o indivíduo for heterozigoto (A / a ou a / A), metade dos gametas carregará um alelo (A) e a outra metade o outro (a). Quando a reprodução sexual é concluída, um novo zigoto é formado, os gametas masculino e feminino se fundem para formar uma nova célula diplóide e um novo par de cromossomos e, portanto, alelos são estabelecidos..
Este processo origina um novo genótipo que é determinado pelos alelos fornecidos pelo gameta masculino e pelo gameta feminino..
Na genética mendeliana, os fenótipos homozigotos e heterozigotos não têm a mesma probabilidade de aparecer em uma população, entretanto, as possíveis combinações alélicas associadas aos fenótipos podem ser inferidas ou determinadas por meio de análise cruzada genética..
Se ambos os pais forem homozigotos para um gene do tipo dominante (A / A), então os gametas de ambos serão do tipo A em sua totalidade e sua união resultará invariavelmente em um genótipo A / A..
Se ambos os pais têm um genótipo homozigoto recessivo (a / a), então a prole invariavelmente resultará em um genótipo homozigoto recessivo também..
Na teoria da evolução, é dito que o motor da evolução é a mudança e no nível genético a mudança ocorre por meio de mutações e recombinações..
As mutações geralmente envolvem mudanças em alguma base de nucleotídeo de um gene, embora possam ser de mais de uma base.
A maioria das mutações são consideradas eventos espontâneos associados à taxa de erro ou fidelidade das polimerases durante a transcrição e replicação do DNA..
Também há muitas evidências de fenômenos físicos que causam mutações no nível genético. Por sua vez, as recombinações podem produzir trocas de seções inteiras de cromossomos, mas estão associadas apenas a eventos de duplicação celular, como mitose e meiose..
Na verdade, eles são considerados um mecanismo básico para gerar variabilidade genotípica durante a formação de gametas. A incorporação da variabilidade genética é uma marca registrada da reprodução sexual.
Focado nos genes, atualmente se considera que o motor da herança e, portanto, da evolução, são genes que apresentam mais de um alelo..
Esses genes que têm apenas um alelo dificilmente podem causar uma mudança evolutiva se todos os indivíduos na população tiverem duas cópias do mesmo alelo, conforme exemplificado acima..
Isso ocorre porque, quando a informação genética é passada de uma geração para outra, dificilmente serão encontradas mudanças nessa população, a menos que existam forças que produzam variações nos genes, como também os mencionados acima..
Os modelos evolutivos mais simples são aqueles que consideram apenas um locus e seu objetivo é tentar prever as frequências dos genótipos na próxima geração, a partir dos dados da geração existente..


Ainda sem comentários