O miofilamentos são as proteínas contráteis das miofibrilas, que são as unidades estruturais das células musculares, células alongadas que são chamadas de fibras musculares.
As fibras musculares e seus componentes têm nomes específicos. Por exemplo, a membrana, o citoplasma, a mitocôndria e o retículo endoplasmático são conhecidos como sarcolema, sarcoplasma, sarcossomas e retículo sarcoplasmático, respectivamente..
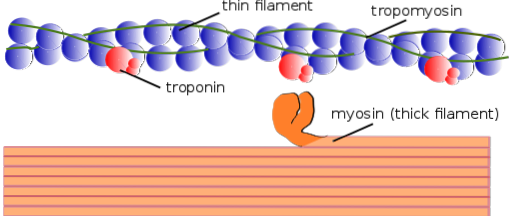
Da mesma forma, os elementos contráteis internos são chamados conjuntamente de miofibrilas; e as proteínas contráteis que compõem as miofibrilas são chamadas de miofilamentos.
Existem dois tipos de miofilamentos: finos e grossos. Os filamentos finos são compostos principalmente de três proteínas: F-actina, tropomiosina e troponina. Os filamentos grossos, por sua vez, são constituídos exclusivamente por outra proteína conhecida como miosina II.
Além destas, existem outras proteínas associadas a filamentos grossos e finos, mas estas não têm funções contráteis, mas sim estruturais, entre as quais estão, para citar algumas, a titina e a nebulina..
Índice do artigo
O arranjo particular dos miofilamentos que compõem as miofibrilas dá origem a dois tipos de fibras musculares: fibras musculares estriadas e fibras musculares lisas.
As fibras musculares estriadas, quando examinadas ao microscópio óptico, apresentam um padrão de estrias ou faixas transversais que se repetem em toda a sua superfície e que dão o nome, ao músculo que as contém, de músculo estriado. Existem dois tipos de fibras musculares estriadas, esqueléticas e cardíacas.
As fibras musculares que não apresentam esse padrão de bandas transversais são chamadas de fibras lisas. São aqueles que constituem os músculos das paredes vasculares e das vísceras.
Esses miofilamentos são compostos por F actina e duas proteínas associadas: a tropomiosina e a troponina, que possuem funções regulatórias..
F actina, ou actina filamentosa, é um polímero de outra proteína globular menor chamada G actina ou actina globular, de cerca de 42 kDa de peso molecular. Possui um sítio de ligação para a miosina e é organizado formando duas cadeias dispostas como uma dupla hélice composta de aproximadamente 13 monômeros por volta..
Os filamentos de F-actina são caracterizados por apresentarem dois pólos: um positivo, direcionado para o disco Z, e o outro negativo, disposto em direção ao centro do sarcômero.
A tropomiosina também é composta por uma dupla cadeia polipeptídica de dupla hélice. É uma proteína de 64 kDa que forma filamentos que estão localizados nas ranhuras deixadas pelas cadeias de dupla hélice dos finos filamentos de F-actina, como se “preenchendo” os espaços vazios da hélice..
Em repouso, a tropomiosina cobre ou "cobre" os locais de ligação da actina para a miosina, impedindo a interação de ambas as proteínas, que é o que causa a contração muscular. Em torno de cada filamento fino e cerca de 25-30m do início de cada tropomiosina está outra proteína chamada troponina.
A troponina (Tn) é um complexo proteico composto de três subunidades polipeptídicas globulares chamadas troponina T, C e I. Cada molécula de troponina tem um complexo de troponina associado que a regula e, juntas, são responsáveis pela regulação da iniciação e terminação da contração muscular..
Os filamentos grossos são polímeros de miosina II, pesando 510 kDa e consistindo em duas cadeias pesadas de 222 kDa cada e quatro cadeias leves. As cadeias leves são de dois tipos: cadeias leves essenciais de 18 kDa e cadeias leves regulatórias de 22 kDa..
Cada cadeia pesada da miosina II tem a forma de uma haste com uma pequena cabeça globular em sua extremidade que se projeta quase 90⁰ e tem dois sítios de ligação, um para a actina e outro para o ATP. É por isso que essas proteínas pertencem à família ATPase..
Um filamento espesso é composto por mais de 200 moléculas de miosina II. A cabeça globular de cada uma dessas moléculas atua como uma “pá” durante a contração, empurrando a actina à qual está fixada para que deslize em direção ao centro do sarcômero..
Em uma fibra muscular estriada esquelética, as miofibrilas ocupam a maior parte do sarcoplasma e são organizadas em conjuntos longitudinais e ordenados por toda a célula.
Em um corte longitudinal visto ao microscópio óptico, são observadas bandas claras, denominadas Bandas I, e bandas escuras, denominadas Bandas A. Essas bandas correspondem ao arranjo ordenado das miofibrilas e, portanto, dos miofilamentos que as compõem..
No centro da Banda I há uma linha escura e fina chamada Linha ou Disco Z. O centro de cada Banda A tem uma área mais clara conhecida como Banda H, que é dividida centralmente por uma linha mais escura chamada Linha M.
Delimitado entre duas linhas Z, é descrita uma estrutura chamada sarcômero, que é a unidade funcional do músculo esquelético. Um sarcômero é composto de miofilamentos contráteis dispostos de maneira ordenada nas bandas A, H e uma hemi-banda I em cada extremidade..
As bandas I contêm apenas filamentos finos, a banda A contém filamentos grossos entrelaçados em suas duas extremidades com filamentos finos e a banda H contém apenas filamentos grossos.
Ambos os miofilamentos grossos e finos podem ser vistos ao examinar uma amostra de músculo esquelético sob um microscópio eletrônico. Diz-se que estes se "interdigitam" ou "se entrelaçam" uns com os outros em um arranjo sequencial, ordenado e paralelo..
Os filamentos finos se originam dos discos Z e se estendem em ambos os lados na direção oposta e em direção ao centro de cada sarcômero adjacente. Dos discos Z em cada extremidade do sarcômero, no músculo relaxado, a actina viaja para o início da banda H em cada lado.
Assim, nas fibras musculares do músculo esquelético relaxado, os miofilamentos espessos ocupam a área central que forma as faixas escuras ou faixas A; e os filamentos finos se estendem em ambos os lados do sarcômero sem atingir o centro do sarcômero.
Em uma seção transversal na região onde os filamentos grossos e finos se sobrepõem, pode-se observar um padrão hexagonal que inclui o filamento grosso no centro e seis filamentos finos que o circundam, e que estão localizados em cada uma das bordas do hexágono.
Essa organização dos miofilamentos no sarcômero é preservada pela função de uma série de proteínas associadas aos miofilamentos e que possuem funções estruturais, entre as quais se destacam titina, alfa actina, nebulina, mioomesina e proteína C..
Quando a acetilcolina (um neurotransmissor) é liberada na placa neuromuscular pela estimulação de um neurônio motor, a fibra muscular é excitada e os canais de cálcio dependentes de voltagem do retículo sarcoplasmático se abrem.
O cálcio se liga à troponina C, e isso causa uma alteração conformacional na tropomiosina, que expõe os sítios ativos da actina, iniciando a contração. Quando os níveis de cálcio caem, a tropomiosina retorna à sua posição inicial e a contração cessa..
A exposição dos locais de ligação da actina à miosina permite que ambas as proteínas se liguem e a miosina empurre a actina em direção ao centro do sarcômero, deslizando sobre a miosina.
Durante a contração muscular, as linhas Z de cada sarcômero se aproximam do centro, aproximando-se da linha M, aumentando a interdigitação entre a actina e a miosina e reduzindo o tamanho das bandas I e H. O grau de encurtamento dependerá da soma do encurtamento de cada um dos sarcômeros do músculo contraído.



Ainda sem comentários