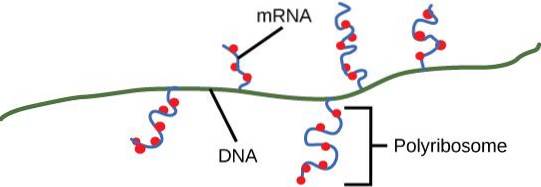
UMA polissomo é um grupo de ribossomos recrutados para a tradução do mesmo RNA mensageiro (mRNA). A estrutura é mais conhecida como polirribossomo, ou ergossoma menos comum..
Os polissomos permitem o aumento da produção de proteínas daqueles mensageiros que são submetidos à tradução simultânea por vários ribossomos. Os polissomos também participam de processos de dobramento co-traducionais e na aquisição de estruturas quaternárias por proteínas recém-sintetizadas..
Os polissomos, junto com os chamados corpos P e grânulos de estresse, controlam o destino e a função dos mensageiros nas células eucarióticas..
Polissomos foram observados em células procarióticas e eucarióticas. Isso significa que esse tipo de formação macromolecular tem uma longa história no mundo celular. Um polissomo pode ser composto de pelo menos dois ribossomos no mesmo mensageiro, mas geralmente há mais de dois.
Em pelo menos uma célula de mamífero, podem existir até 10.000.000 de ribossomos. Muitos foram observados como livres, mas uma grande parte está associada aos polissomos bem conhecidos.
Índice do artigo
Os ribossomos de todos os seres vivos consistem em duas subunidades: a subunidade pequena e a subunidade grande. A pequena subunidade dos ribossomos é responsável pela leitura do RNA mensageiro.
A grande subunidade é responsável pela adição linear de aminoácidos ao peptídeo nascente. Uma unidade de tradução ativa é aquela em que um mRNA foi capaz de recrutar e permitir a montagem do ribossomo. Depois disso, a leitura dos tripletos no mensageiro e a interação com o tRNA carregado correspondente prossegue sequencialmente..
Os ribossomos são os blocos de construção dos polissomos. Na verdade, as duas formas de traduzir um mensageiro podem coexistir na mesma célula. Se todos os componentes que compõem o mecanismo de translação da célula forem purificados, encontraremos quatro frações principais:
Em células eucarióticas, os mRNAs são exportados do núcleo como ribonucleoproteínas mensageiras. Ou seja, o mensageiro é acoplado a várias proteínas que irão determinar sua exportação, mobilização e tradução..
Dentre eles, há vários que interagem com a proteína PABP ligada à cauda poliA 3 'do mensageiro. Outros, como o complexo CBP20 / CBP80, irão se ligar à capa 5 'do mRNA.
A liberação do complexo CBP20 / CBP80 e o recrutamento de subunidades ribossômicas na capa 5 'definem a formação do ribossomo.
A tradução começa e novos ribossomos são montados no capô de 5 '. Isso acontece por um número limitado de vezes que depende de cada mensageiro e do tipo de polissomo em questão..
Após esta etapa, os fatores de alongamento da tradução associados ao cap na extremidade 5 'interagem com a proteína PABP ligada à extremidade 3' do mRNA. Um círculo é então formado definido pela união das regiões não traduzíveis do mensageiro. Assim, tantos ribossomos são recrutados quanto o comprimento do mensageiro, e outros fatores, permitem.

Outros polissomos podem adotar uma configuração linear dupla linha ou espiral com quatro ribossomos por volta. A forma circular tem sido mais fortemente associada a polissomos livres.
Os polissomos são formados em unidades de tradução ativas (inicialmente monossomos) com a adição sequencial de outros ribossomos no mesmo mRNA.
Dependendo de sua localização subcelular, encontramos três tipos diferentes de polissomos, cada um com suas próprias funções particulares..
São encontrados livres no citoplasma, sem associações aparentes com outras estruturas. Esses polissomos traduzem os mRNAs que codificam para proteínas citosólicas.
Como o envelope nuclear é uma extensão do retículo endoplasmático, este tipo de polissoma também pode ser associado ao envelope nuclear externo.
Nesses polissomos, os mRNAs que codificam dois grupos importantes de proteínas são traduzidos. Alguns, que são uma parte estrutural do retículo endoplasmático ou do complexo de Golgi. Outros, que devem ser modificados pós-tradução e / ou realocados intracelularmente por essas organelas.
Polissomos associados ao citoesqueleto traduzem proteínas de mRNAs que são assimetricamente concentrados em certos compartimentos subcelulares.
Ou seja, ao deixar o núcleo, algumas ribonucleoproteínas mensageiras são mobilizadas para o local onde é necessário o produto que codificam. Essa mobilização é realizada pelo citoesqueleto com a participação de proteínas que se ligam à cauda poliA do mRNA..
Em outras palavras, o citoesqueleto distribui os mensageiros por destino. Esse destino é indicado pela função da proteína e pelo local onde ela deve residir ou agir..
Mesmo que um mRNA seja transcrito, isso não significa necessariamente que ele deva ser traduzido. Se este mRNA é degradado especificamente no citoplasma da célula, a expressão de seu gene é considerada regulada pós-transcricionalmente..
Existem várias maneiras de se conseguir isso, e uma delas é por meio da ação dos chamados genes MIR. O produto final da transcrição de um gene MIR é um microRNA (miRNA).
Estes são complementares ou parcialmente complementares a outros mensageiros cuja tradução eles regulam (silenciamento pós-transcricional). O silenciamento também pode envolver degradação específica de um determinado mensageiro.
Tudo relacionado à tradução, sua compartimentação, regulação e silenciamento gênico pós-transcricional é controlado por polissomos..
Para fazer isso, eles interagem com outras macroestruturas moleculares da célula conhecidas como corpos P e grânulos de estresse. Esses três corpos, mRNAs e microRNAs, definem o proteoma presente em uma célula em um determinado momento..



Ainda sem comentários