As Proteínas G ou proteínas de ligação de nucleotídeo guanina, são proteínas associadas à membrana plasmática pertencentes a uma família de proteínas "acopladoras" de sinal que têm funções importantes em muitos processos de transdução de sinal em organismos eucarióticos..
Na literatura, as proteínas G são descritas como interruptores moleculares binários, uma vez que sua atividade biológica é determinada pelas mudanças em sua estrutura dadas pelas espécies de nucleotídeos às quais são capazes de se ligar: nucleotídeos de guanosina (difosfato (GDP) e trifosfato (GTP) ).
Eles são geralmente ativados por receptores de uma família de proteínas conhecidas como receptores acoplados à proteína G (GPCRs).Receptores acoplados à proteína G "), que recebem um sinal inicial externo e o convertem em mudanças conformacionais que desencadeiam a ativação, que posteriormente se traduz na ativação de outra proteína efetora.
Alguns autores consideram que os genes codificadores desta família de proteínas evoluíram por duplicação e divergência de um gene ancestral comum, cujo produto foi cada vez mais refinado e especializado..
Entre a grande variedade de funções celulares que essas proteínas apresentam, estão a translocação de macromoléculas durante a síntese protéica, a transdução de sinais hormonais e estímulos sensoriais, bem como a regulação da proliferação e diferenciação celular..
Duas classes desse tipo de proteína foram descritas: proteínas G pequenas e proteínas G heterotriméricas. A primeira estrutura tridimensional de uma proteína G foi obtida há mais de uma década a partir de uma pequena proteína G conhecida como Ras.
Índice do artigo
Estruturalmente falando, dois tipos de proteínas G são reconhecidos: as proteínas G pequenas e as proteínas G heterotriméricas, muito mais complexas..
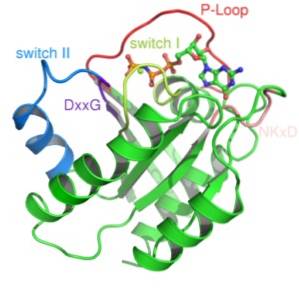
Proteínas G pequenas são constituídas por um único polipeptídeo de cerca de 200 resíduos de aminoácidos e cerca de 20-40 kDa, e em sua estrutura há um domínio catalítico conservado (o domínio G) composto por cinco hélices α, seis folhas dobradas β , e cinco loops polipeptídicos.
As proteínas G heterotriméricas, por outro lado, são proteínas integrais de membrana compostas por três cadeias polipeptídicas, conhecidas como subunidades α, β e γ..
-A subunidade α pesa entre 40 e 52 kDa, tem uma região de ligação do nucleotídeo guanina e tem atividade GTPase para hidrolisar as ligações entre os grupos fosfato do GTP..
As subunidades α de diferentes proteínas G compartilham alguns domínios estruturais, como aqueles para ligação e hidrólise de GTP, mas são muito diferentes em seus locais de ligação para o receptor e proteínas efetoras..
-A subunidade β tem um peso molecular ligeiramente inferior (entre 35 e 36 kDa).
-A subunidade γ, por outro lado, é muito menor e tem um peso molecular aproximado de 8 kDa..
Todas as proteínas G heterotriméricas têm 7 domínios transmembranares e compartilham similaridade de sequência com os domínios β e γ. Esses dois domínios estão tão fortemente associados que são vistos como uma única unidade funcional..
Conforme mencionado acima, existem dois tipos de proteínas G: pequenas e heterotriméricas..
Proteínas G pequenas têm papéis no crescimento celular, secreção de proteínas e interação de vesículas intracelulares. Por sua vez, as proteínas G heterotriméricas estão associadas à transdução de sinais de receptores de superfície e também atuam como interruptores que alternam entre dois estados dependendo do nucleotídeo associado..
Essas proteínas também são chamadas de pequenas GTPases, pequenas proteínas de ligação a GTP ou superfamília de proteínas Ras e formam uma superfamília independente dentro da grande classe de GTP hidrolases com funções regulatórias..
Essas proteínas são muito diversas e controlam vários processos celulares. Eles são caracterizados por um domínio de ligação a GTP conservado, o domínio "G". A ligação desse nucleotídeo fosfato causa mudanças conformacionais importantes em seu domínio catalítico em proteínas G pequenas..
Sua atividade está intimamente relacionada às proteínas ativadoras de GTPase (GAP).Proteína ativadora de GTPase ") e com fatores de troca de nucleotídeo guanina (GEF)Fator de troca de nucleotídeo de guanina ”).
Cinco classes ou famílias de pequenas proteínas G foram descritas em eucariotos:
-Ras
-Rho
-Rab
-Sar1 / Arf
-Correu
As proteínas Ras e Rho controlam a expressão gênica e as proteínas Rho também modulam a reorganização do citoesqueleto. As proteínas do grupo Rab e Sar1 / Arf influenciam o transporte vesicular e as proteínas Ran regulam o transporte nuclear e o ciclo celular..
Esse tipo de proteína também merece associação com dois outros fatores protéicos, de forma que a via de sinalização do ambiente externo para o interior da célula seja composta por três elementos na seguinte ordem:
Existe uma grande diversidade de proteínas G heterotriméricas e isso está relacionado à grande diversidade de subunidades α que existem na natureza, nas quais apenas 20% da sequência de aminoácidos é conservada..
Normalmente, as proteínas G heterotriméricas são identificadas graças à diversidade da subunidade α, baseada principalmente em suas semelhanças funcionais e de sequência..
As subunidades α são compostas por quatro famílias (a família Gs, a família Gi / o, a família Gq e a família G12). Cada família é composta por um “isótipo” diferente que, juntos, adiciona mais de 15 formas diferentes de subunidades α..
Esta família contém representantes que também participam da regulação positiva das proteínas adenilato ciclase e é expressa na maioria dos tipos de células. É composto por dois membros: Gs e Golf.
O subscrito "s " refere-se a estimulação (do inglês “estimulação ") e o subscrito “olf " refere-se a "cheiro" (do inglês "Olfato") As proteínas do golfe são expressas especialmente em neurônios sensoriais responsáveis pelo cheiro.
Esta é a maior e mais diversa família. Eles são expressos em muitos tipos de células e medeiam a inibição dependente do receptor de vários tipos de adenil ciclases (o subscrito “eu " refere-se a inibição).
As proteínas com as subunidades α do grupo Go são expressas especialmente em células do sistema nervoso central e têm duas variantes: A e B..
As proteínas com esta família de subunidades α são responsáveis pela regulação da fosfolipase C. Esta família consiste em quatro membros cujas subunidades α são expressas por genes diferentes. Eles são abundantes nas células do fígado, células renais e pulmões.
Esta família é expressa de forma ubíqua nos organismos e não se sabe ao certo quais são exatamente os processos celulares regulados por proteínas com essas subunidades..
Subunidades Β e γ
Embora a diversidade de estruturas alfa seja decisiva para a identificação de proteínas heterotriméricas, também há muita diversidade em relação às outras duas subunidades: beta e gama..
As proteínas G participam da "canalização" de sinais de receptores na membrana plasmática para canais ou enzimas efetoras..
O exemplo mais comum da função desse tipo de proteína está na regulação da enzima adenilato ciclase, enzima responsável pela síntese da adenosina 3 ', 5'-monofosfato ou simplesmente AMP cíclico, molécula que tem importantes funções como segundo mensageiro em muitos processos celulares conhecidos:
-Fosforilação seletiva de proteínas com funções específicas
-Transcrição genética
-Reorganização do citoesqueleto
-Secreção
-Despolarização da membrana
Também participam indiretamente da regulação da cascata de sinalização do inositol (fosfatidilinositol e seus derivados fosforilados), responsáveis pelo controle de processos cálcio-dependentes, como a quimiotaxia e a secreção de fatores solúveis..
Muitos canais iônicos e proteínas de transporte são controlados diretamente por proteínas da família das proteínas G. Da mesma forma, essas proteínas estão envolvidas em diversos processos sensoriais como visão, olfato, entre outros..
O modo de interação de uma proteína G com as proteínas efetoras é específico para cada classe ou família de proteínas.
Para proteínas G acopladas a receptores de membrana (proteínas G heterotriméricas), a ligação de um nucleotídeo de guanina, como GDP ou difosfato de guanosina à subunidade α, causa a associação das três subunidades, formando um complexo conhecido como Gαβγ ou G-GDP, que é preso à membrana.
Se a molécula de GDP for posteriormente trocada por uma molécula de GTP, a subunidade α ligada ao GTP se dissocia das subunidades β e γ, formando um complexo separado conhecido como Gα-GTP, que é capaz de alterar a atividade de suas enzimas ou proteínas transportadoras alvo.
A atividade hidrolítica dessa subunidade permite que ela encerre a ativação, trocando o GTP por um novo GDP, passando para a conformação inativa..
Na ausência de receptores excitados que se associam às proteínas G, esse processo de troca de GDP por GTP é muito lento, o que significa que proteínas G heterotriméricas apenas trocam GDP por GTP a uma taxa fisiologicamente significativa quando estão ligadas a seus receptores excitados..



Ainda sem comentários