O epistasia, em genética, é o estudo da interação entre os diferentes genes que codificam para o mesmo personagem. Ou seja, é a manifestação de uma característica que surge da interação entre alelos de genes em diferentes loci.
Quando falamos sobre as relações que estabelecem alelos do mesmo gene, nos referimos a relações alélicas. Ou seja, para alelos do mesmo locus ou alelos alelomórficos. Essas são as interações bem conhecidas de dominância completa, dominância incompleta, codominância e letalidade entre alelos do mesmo gene..
Nas relações entre alelos de diferentes loci, ao contrário, falamos de alelos não alelomórficos. Essas são as chamadas interações gênicas, que de alguma forma são todas epistáticas.
A epistasia permite analisar se a expressão de um gene determina a expressão de outro. Em tal caso, tal gene seria epistático no segundo; o segundo seria hipostático no primeiro. A análise da epistasia também permite determinar a ordem em que atuam os genes que definem o mesmo fenótipo..
A epistasia mais simples analisa como dois genes diferentes interagem para conferir o mesmo fenótipo. Mas, obviamente, pode haver muitos mais genes.
Para a análise da epistasia simples basear-nos-emos nas variações das proporções dos cruzamentos diíbridos clássicos. Ou seja, para as modificações da proporção 9: 3: 3: 1, e para ela.
Índice do artigo
Essa proporção surge da combinação da análise de herança de dois caracteres diferentes. Ou seja, é o produto da combinação de duas segregações fenotípicas independentes (3: 1) X (3: 1).
Quando Mendel analisou, por exemplo, estatura da planta ou cor da semente, cada personagem segregou 3 para 1. Quando ele os analisou juntos, mesmo que fossem dois personagens diferentes, cada um segregou 3 para 1. Ou seja, eles foram distribuídos independentemente.
No entanto, quando Mendel analisou os caracteres em pares, eles resultaram nas conhecidas classes fenotípicas 9, 3, 3 e 1. Mas essas classes eram somas de dois caracteres. diferente. E nunca, nenhum personagem afetou como o outro se manifestou.
O anterior foi a explicação da proporção mendeliana clássica. Portanto, não é um caso de epistasia. Epistasia estuda os casos de herança do mesmo caráter determinado por vários genes.
O caso anterior, ou segunda lei de Mendel, era de herança de dois personagens diferentes. Aquelas explicadas posteriormente são verdadeiras proporções epistáticas e envolvem apenas alelos não alelomórficos..
Esse caso é encontrado quando o mesmo personagem apresenta quatro diferentes manifestações fenotípicas na proporção de 9: 3: 3: 1. Portanto, não pode ser uma interação alélica (monogênica) como aquela que leva ao aparecimento de quatro grupos sanguíneos diferentes no sistema ABO..
Tomemos como exemplo o cruzamento entre um indivíduo heterozigoto com sangue tipo A e um indivíduo heterozigoto com sangue tipo B. euPARAeu X euBeu. Isso nos daria uma proporção de 1: 1: 1: 1 de indivíduos euPARAeu (Tipo A), euPARAeuB (Tipo AB), euBeu (Tipo B) e ii (Tipo O).
Ao contrário, uma verdadeira relação epistática dupla dominante (9: 3: 3: 1) é observada na forma de crista de galo. Existem quatro classes fenotípicas, mas em uma proporção de 9: 3: 3: 1.
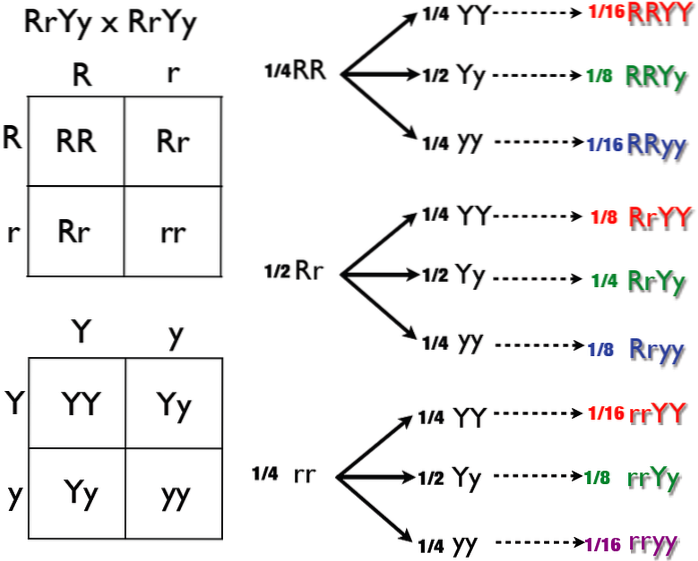
Dois genes participam de sua determinação e manifestação, vamos chamá-los R Y P. Independentemente disso, os alelos R Y P mostrar domínio completo sobre alelos r Y p, respectivamente.
Da travessia RrPp X RrPp podemos obter as classes fenotípicas 9 R_P_, 3 R_pp, 3 rrP_ e 1 rrpp. O símbolo "_" significa que o alelo pode ser dominante ou recessivo. O fenótipo associado permanece o mesmo.
A classe 9 R_P_ é representada por galos com crista de nogueira, 3 R_pp por galos com crista de rosa. Galos com crista de ervilha seriam Classe 3 rrP_; aqueles da classe rrpp têm uma crista simples.

Na epistasia dupla dominante, cada classe 3 surge do efeito de dominância do gene R ou P. A classe 9 é representada por aquele em que se manifestam os alelos dominantes R e P. Finalmente, na classe 1 rrpp, os alelos estão ausentes. de ambos os genes.
Nessa interação epistática, um gene não suprime a manifestação de outro. Pelo contrário, ambos os genes codificam para a manifestação do mesmo traço, mas sem efeito aditivo..
Portanto, a presença de pelo menos um alelo dominante de qualquer um dos dois genes de loci diferentes permite a manifestação do traço na classe 15. A ausência de alelos dominantes (a classe dupla recessiva) determina o fenótipo da classe 1.
Os produtos dos genes participam da manifestação da cor do grão de trigo PARA Eu B. Ou seja, qualquer um desses produtos (ou ambos) pode levar à reação bioquímica que transforma o precursor em pigmento..
A única classe que não produz nenhum deles é a classe 1 aabb. Portanto, as classes 9 A_B_, 3 A_bb e 3 aaB_ irão produzir grãos pigmentados, e a minoria restante não irá.
Aqui encontramos um caso de deleção dominante de um gene (hipostático) devido à presença de pelo menos um alelo dominante do outro (epistático). Ou seja, formalmente falando, um gene suprime a ação do outro..
Se for a supressão dominante de D sobre K, teríamos o mesmo fenótipo associado às classes 9 D_K_, 3 D_kk e 1 ddkk. Classe 3 ddK_ seria a única mostrando o traço não suprimido.
A classe recessiva dupla é adicionada às classes 9 D_K_ e 3 D_kk porque não produz o que o gene hipostático K codifica. Não porque seja suprimida por D, que em qualquer caso também não está lá, mas porque não produz K.
Essa proporção também é chamada de epistasia dominante e recessiva. O dominante é o de K sobre D / d. A epistasia recessiva seria a de dd sobre K / k.
Por exemplo, flores de prímula devem sua cor à manifestação de dois genes. The Gen K que codifica para a produção do pigmento malvidina, e o gene D quais códigos para a exclusão de malvidin.
Apenas plantas ddKK ou ddKk (ou seja, classe 3 ddK_) produzirá malvidina e será de cor azul. Qualquer outro genótipo dará origem a plantas com flores turquesa.
Nesse caso, a presença de pelo menos um alelo dominante de cada gene do par é necessária para que o personagem se manifeste. Digamos que sejam os genes C Y P. Ou seja, o estado homozigoto recessivo de um dos genes do par (DC ou pp) impossibilita a manifestação do personagem.
Em outras palavras, apenas a classe 9 C_P_ tem pelo menos um alelo dominante C e um alelo dominante P. Para que a característica se manifeste, os produtos funcionais dos dois genes devem estar presentes.
Essa interação é epistática porque a falta de expressão de um gene impede que o outro gene se manifeste. É duplo, porque o inverso também é verdadeiro.
Um exemplo clássico que ilustra esse caso é o das flores de ervilha. As plantas CCpp e as plantas ccPP eles têm flores brancas. Os híbridos CcPp dos cruzamentos entre eles apresentam flores roxas.
Se duas dessas plantas di-híbridas forem cruzadas, obteremos a classe 9 C-P_, que terá flores roxas. As classes 3 C_pp, 3 ccP_ e ccpp serão flores brancas.

Da proporção proposta na segunda lei de Mendel, temos outros casos adicionais que merecem destaque..
Chamamos a relação modificada de 9: 4: 3 de epistasia recessiva por um bom motivo. Quando um gene é homozigoto para o gene recessivo, ele evita a expressão do outro gene - mesmo que seja dominante.
Tomemos por exemplo a epistasia recessiva do genótipo aa sobre aqueles do gene B. A classe 9 é a já reconhecida 9 A_B_. Para a classe 4, à classe 1 aabb, devem ser adicionados os da classe 3 aaB_, com o mesmo fenótipo. Classe 3 seriam aqueles da classe 3 A_bb.
Na interação epistática de genes duplicados, a proporção fenotípica observada é de 9: 6: 1. Todos os indivíduos da classe 9 A_B_ têm pelo menos um alelo de cada gene PARA ou B. Todos eles têm o mesmo fenótipo.
Pelo contrário, nas classes 3 A_bb e 3 aaBb, existem apenas alelos dominantes de A ou B. Neste caso, também existe um único e o mesmo fenótipo - mas diferente dos outros. Finalmente, na classe 1 aabb nenhum alelo dominante de qualquer um dos genes está presente e representa outro fenótipo.
Talvez a classe mais confusa seja a epistasia dominante, que mostra a proporção fenotípica de 12: 3: 1. Aqui, o domínio de A (epistático) sobre B (hipostático) faz com que a classe 9 A-B_ junte-se à classe 3 A_bb.
O fenótipo de B só se manifestará quando A não estiver presente na classe 3 aaB_. O aabb duplo recessivo classe 1 não se manifestará nem o fenótipo associado ao gene A / a nem com o gene B / b.
Outras proporções fenotípicas epistáticas que não têm um nome específico são 7: 6: 3, 3: 6: 3: 4 e 11: 5.


Ainda sem comentários