
O teoria de dobramento de membrana propõe que as membranas das organelas se originaram da expansão e invaginação da membrana plasmática. J.D Robertson, um pioneiro em microscopia eletrônica, observou em 1962 que vários corpos intracelulares possuíam uma estrutura que era visualmente idêntica à membrana plasmática..
A ideia de uma estrutura que delimite as células surgiu imediatamente após o surgimento do conceito de "célula", para o qual foram realizados vários estudos para elucidar as características dessa estrutura..
Índice do artigo
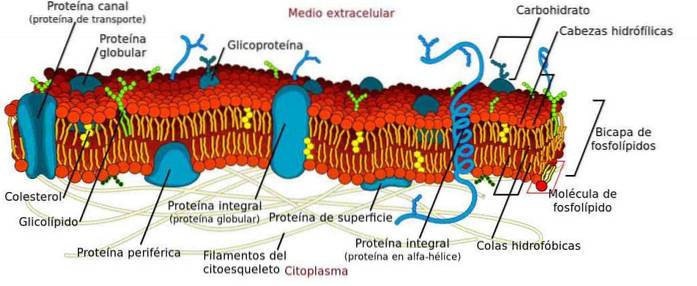
A membrana plasmática é uma estrutura formada por uma dupla camada de fosfolipídios organizada de forma que os grupos polares estejam orientados para o citosol e o meio extracelular, enquanto os grupos apolares se organizem para o interior da membrana..
Sua principal função é definir células, tanto eucarióticas quanto procarióticas, uma vez que separa fisicamente o citoplasma do meio extracelular..
Apesar de sua função estrutural, sabe-se que a membrana não é estática, mas sim uma barreira elástica e dinâmica onde ocorre um grande número de processos essenciais para a célula..
Alguns processos que ocorrem na membrana são a ancoragem do citoesqueleto, o transporte de moléculas, a sinalização e a conexão com outras células para formar os tecidos. Além disso, uma grande variedade de organelas também possui uma membrana na qual ocorrem outros processos de grande importância..
Muito antes de Robertson propor a teoria do dobramento da membrana em 1962, os primeiros estudos foram realizados para determinar a aparência dessa estrutura. Na ausência do microscópio eletrônico, predominaram os estudos eletrofisiológicos, entre os quais:
Overton observou que os lipídios atravessam a membrana celular com mais facilidade do que moléculas de outra natureza, portanto, inferiu que a membrana deve ser composta, em sua maioria, por lipídios..
J. Bernstein apresentou sua hipótese, que mencionava que as células consistiam em uma solução com íons livres delimitados por uma fina camada impermeável às referidas moléculas carregadas..
Fricke mediu a capacidade da membrana do eritrócito de armazenar cargas (capacitância), determinando que esse valor era de 0,81 µF / cmdois.
Posteriormente foi determinado que as membranas de outros tipos de células tinham valores de capacitância semelhantes, portanto, a membrana deveria ser uma estrutura unitária..
Gorter e Grendel mediram a área dos eritrócitos de mamíferos com a ajuda de um microscópio. Eles então extraíram lipídios de um número conhecido desse tipo de célula e mediram a área que ocupavam..
Como resultado, eles obtiveram uma relação célula: membrana de 1: 2. Isso significava que a membrana celular tinha uma estrutura dupla, dando origem ao termo "bicamada lipídica".
Estudos anteriores a 1935 sugeriam a presença de proteínas na membrana, o que levou Danielli e Davson a propor o modelo Sandwich ou o modelo Proteína-Lípido-Proteína..
Segundo esse modelo, a membrana plasmática consiste em duas camadas de fosfolipídios que se encontram entre duas camadas de proteínas, que se associam à membrana por meio de interações eletrostáticas..
Em 1959, graças ao surgimento da microscopia eletrônica, J. David Robertson coletou evidências suficientes para confirmar e complementar os modelos propostos por Gorter e Grendel (1925) e Danielli e Davson (1935), e propor o modelo de "Membrana Unitária"..
Esse modelo mantém a característica do modelo proposto por Danielli e Davson de bicamada lipídica, com a variação da camada protéica que, neste caso, é assimétrica e descontínua. .
A chegada da microscopia eletrônica permitiu ter uma ideia bastante clara sobre como a membrana plasmática foi formada..
No entanto, esse fato foi acompanhado pela visualização de múltiplas membranas intracitoplasmáticas que formaram compartimentos intracelulares, o que levou Robertson em 1962 a propor a "Teoria do Dobramento da Membrana".
A teoria do dobramento da membrana consiste em que a membrana plasmática aumenta sua superfície e invagina para dar origem às membranas intracitoplasmáticas, essas membranas circundam as moléculas que estavam no citosol, originando assim as organelas..
Segundo essa teoria, o envelope nuclear, o retículo endoplasmático, o aparelho de Golgi, os lisossomos e os vacúolos poderiam ter se originado dessa forma..
A continuidade que existe entre a membrana plasmática e as três primeiras organelas mencionadas acima foi confirmada por estudos de microscopia eletrônica em diferentes tipos de células..
No entanto, Robertson também propôs em sua teoria que organelas vesiculares, como lisossomos e vacúolos, também se originaram por invaginações que posteriormente se separaram da membrana..
Pelas características da teoria do dobramento da membrana, é considerada uma extensão do modelo de membrana unitária que ele mesmo propôs em 1959.
As micrografias feitas por Robertson mostram que todas essas membranas são iguais e, portanto, devem ter uma composição bastante semelhante..
No entanto, a especialização das organelas modifica consideravelmente a composição das membranas, o que reduz as propriedades que elas têm em comum a nível bioquímico e molecular..
Da mesma forma, mantém-se o facto de as membranas terem como principal função servir de barreira estável para meios aquosos..
Graças a todos os testes realizados entre 1895 e 1965, especialmente os estudos de microscopia realizados por J.D. Robertson, a importância das membranas celulares foi enfatizada.
A partir do seu modelo unitário, passou a ser destacado o papel essencial que a membrana desempenha na estrutura e função das células, a ponto de o estudo desta estrutura ser considerado uma questão fundamental na biologia atual..
No entanto, com relação à contribuição da teoria do dobramento da membrana, isso não é atualmente aceito. Porém, na época, isso levou mais especialistas na área a tentarem elucidar a origem não só das membranas celulares, mas também a origem da própria célula eucariótica, como fez Lynn Margulis em 1967 ao levantar a teoria endossimbiótica..



Ainda sem comentários